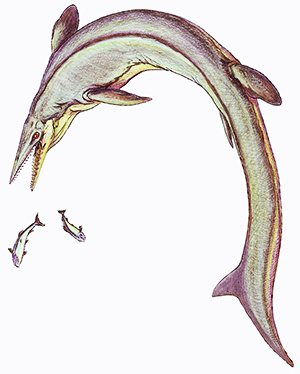
Il genere annovera attualmente le seguenti specie: Clidastes propython Cope, 1869 (nuova specie) - Clidastes liodontus Merriam, 1894
Descrizione
Questo mosasauro era relativamente piccolo se confrontato con la maggior parte dei suoi simili. In media raggiungeva la lunghezza di circa 4 metri, anche se sono stati ritrovati esemplari lunghi anche 6 metri. Il corpo del Clidastes era snello e allungato, mentre la coda appiattita lateralmente era piuttosto corta rispetto a quella di altri mosasauri. Le spine neurali espanse verso la punta della coda indicano che questo animale utilizzava questo strumento per spostarsi in acqua, anche se non è chiaro se il possedesse la coda bilobata come altri mosasauri (ad es. il Platecarpus). Alcune caratteristiche del Clidastes erano primitive: la struttura delle zampe, ad esempio, con poche falangi per dito; le dita, inoltre, non erano compatte fra loro ed era presente un'ampia membrana tra un dito e l'altro. Il carpo, inoltre, era di foggia primitiva, mentre omero, ulna e radio erano altamente modificati, con forme appiattite e allargate. Il cranio era piuttosto mobile (anche se non mobile come quello dei mosasauri plioplatecarpini) e possedeva un muso appuntito e triangolare. I denti erano compressi lateralmente e dotati di doppia carena (anteriore e posteriore), ed erano ricoperti di smalto liscio, molto simili a quelli del Mosasaurus. A quest'ultimo genere il Clidastes assomigliava anche per le vertebre cervicali, che differivano (oltre che per le dimensioni minori) per la forma più allungata. La coda, invece, aveva una forma diversa rispetto a quella del Mosasaurus. Questo mosasauro, come molti altri scoperti sul finire dell'Ottocento, ha una storia tassonomica complicata. Il genere Clidastes è stato descritto per la prima volta da Edward Drinker Cope nel 1868, sulla base di una vertebra isolata ritrovata nella formazione Marshalltown in New Jersey. Cope istituì la specie tipo (Clidastes iguanavus) sulla base di quest'unico resto fossile, riscontrando presunte somiglianze con le vertebre delle odierne iguane. La vertebra possedeva uno zigosfeno e uno zigantro (articolazioni accessorie che aiutano a incastrare le vertebre le une con le altre, comuni in molte lucertole attuali e nei serpenti) e portò Cope alla convinzione che i mosasauri fossero animali serpentiformi. Successive scoperte smentirono questa ricostruzione, ma in ogni caso l'idea di Cope secondo cui mosasauri e serpenti erano strettamente imparentati è ancora valida. Successivamente vennero scoperti numerosi altri fossili di mosasauri che Cope stesso attribuì al genere Clidastes, e la vertebra che costituiva l'esemplare tipo venne sostanzialmente dimenticata. Nel 1992, per conservare il nome generico Clidastes, venne proposto quindi di cambiare la specie tipo e l'olotipo: Clidastes propython, sulla base di uno scheletro quasi completo proveniente dalla formazione Selma dell'Alabama, che Cope descrisse nel 1869 e attribuì a Clidastes per la presenza di zigosfeni e zigantri sulle vertebre. Attualmente, la vertebra di Clidastes iguanavus non è più considerata diagnostica e si suppone addirittura che possa essere appartenuta al genere Mosasaurus. Il Clidastes propython è noto per numerosi esemplari, provenienti principalmente dal Santoniano/Campaniano inferiore del Nordamerica e in particolare dall'Alabama e dal Kansas, ma anche da Texas, Manitoba, Dakota del Sud, Colorado; resti attribuiti a questa specie sono stati ritrovati anche in Svezia (Lindgren e Siverson, 2004) e in Russia (Grigoriev et al., 2015). Oltre al C. propython, è da ricordare il C. liodontus, ben conosciuto grazie a numerosi esemplari; questa specie venne descritta per la prima volta da Merriam nel 1894 e venne avvicinata a un altro mosasauro, il Liodon (da qui il nome specifico), sulla base dei denti lisci e taglienti. Il C. liodontus proviene da terreni più antichi (Coniaciano/Santoniano) rispetto a quelli in cui sono stati ritrovati i fossili di C. propython. Un altro esemplare proveniente dal Mooreville Chalk dell'Alabama, Clidastes moorevillensis, rimasto a lungo un nomen nudum, venne descritto solo nel 2012 (Karl e Nyhuis, 2012). Altri resti attribuiti al Clidastes (principalmente denti e mascelle) sono noti in Europa (Svezia, Germania, Inghilterra), in Nordafrica e in Asia sudoccidentale. I resti più recenti attribuiti al Clidastes provengono dal Campaniano superiore della Germania (Diedrich e Mulder, 2004). Il Clidastes è considerato un rappresentante basale della sottofamiglia dei mosasaurini, comprendente mosasauri dal corpo allungato e dalla coda relativamente corta, con vertebre dotate di zigosfeni e zigantri. Al contrario di forme più specializzate come il Plotosaurus e il Mosasaurus, il Clidastes possedeva ancora un cranio dalle articolazioni relativamente mobili. Secondo uno studio di Bell (1997) il Clidastes è il rappresentante più basale dei mosasaurini. Lo stesso studio conclude che il Clidastes non sarebbe un genere strettamente monofiletico, bensì comprenderebbe una serie di forme successive e sempre più derivate di mosasaurini; secondo questa ipotesi il C. liodontus sarebbe il mosasaurino più basale, mentre il C. propython costituirebbe il sister group dei mosasaurini derivati (come il Globidens, il Mosasaurus e il Plotosaurus). Le spine neurali espanse verso la punta della coda aiutavano l'animale a spostarsi con forza, e suggeriscono che il Clidastes fosse un veloce nuotatore. Uno studio di Russell (1967) ha ipotizzato che le lunghe e sottili mascelle di questo mosasauro fossero adatte a mordere rapidamente le prede; i denti erano adatti a tagliare grossi oggetti in pezzi più piccoli che potevano essere inghiottiti dall'animale quando la mandibola veniva mossa in avanti e indietro. Al contrario di molti grandi mosasauri che probabilmente inghiottivano le loro prede intere, il più piccolo Clidastes sembrerebbe essere stato in grado di ridurre una preda in pezzi tramite i suoi morsi. Lo stile di vita del Clidastes è spesso oggetto di dibattito. Nel 1898 Williston considerò questo mosasauro come un predatore di superficie, e questa idea rimase predominante per un secolo. Uno studio di Martin e Rotschild (1989) mise in luce il fatto che le ossa del Clidastes, al contrario di quelle di altri mosasauri (come il Platecarpus), non portavano i segni di sindrome da decompressione (necrosi vascolare): secondo gli studiosi, questa era un'ulteriore prova del fatto che il Clidastes vivesse nei pressi della riva e non fosse adatto a grandi profondità. Un altro studio (Sheldon, 1997) ha invece evidenziato la microstruttura delle costole del Clidastes, caratterizzate da una densità ossea notevolmente bassa (osteoporosi); in vita, le ossa porose dovevano essere riempite con lipidi, che provvedevano al bilanciamento dell'animale in una vasta gamma di profondità. Sulla base di comparazioni con alcuni vertebrati marini attuali, Sheldon ritenne che il Clidastes fosse un nuotatore di profondità, con un torace costruito in modo tale da diminuire la diffusione di gas all'interno dei vasi sanguigni (così da prevenire la necrosi vascolare). Successivi studi hanno invece fatto pensare a al Clidastes come a un nuotatore di superficie, che viveva in acque basse: i fossili svedesi, in particolare, sono stati ritrovati in una zona che, nel Cretaceo, era costituita da un ambiente costiero (Lindgren e Siverson, 2004).
Diffusione
Rettile marino estinto, appartenente alla famiglia dei mosasauridi. Visse nel Cretaceo superiore (Coniaciano - Campaniano, tra 88 e 80 milioni di anni fa). I suoi resti sono stati rinvenuti principalmente in Nordamerica, ma anche in Europa, in Nordafrica e in Asia sudoccidentale.
Bibliografia
Rapid Evolution of Mosasaurs". oceansofkansas.com. Cope, E.D. 1868. On new species of extinct reptiles. Proceedings of the Academy of Natural Sciences of Philadelphia 20: 181. Wright, K. R. (September 23, 1988). The First Record of Clidastes liodontus (Squamata, Mosasauridae) from the Eastern United States. Journal of Vertebrate Paleontology, 8, 3, 343-34- Russell, Dale. A. (6 November 1967). "Systematics and Morphology of American Mosasaurs" (PDF). Bulletin of the Peabody Museum of Natural History (Yale University). Olivier, R., & Maureen, K. (December 01, 2005). Tooth Replacement in the Late Cretaceous Mosasaur Clidastes. Journal of Herpetology, 39, 4. Luan, X., Walker, C., Dangaria, S., Ito, Y., Druzinsky, R., Jarosius, K., Lesot, H Rieppel, O. (January 01, 2009). The mosasaur tooth attachment apparatus as paradigm for the evolution of the gnathostome periodontium. Evolution & Development, 11, 3. Lindgren, J., & Siverson, M. (January 01, 2004). The first record of the mosasaur Clidastes from Europe and its palaeogeographical implications. Acta Palaeontologica Polonica, 49, 219-234. Caldwell, M.W., & Diedrich, C.G. 2005. Remains of Clidastes Cope, 1868, an unexpected mosasaur in the upper Campanian of NW Germany. (Igitur.) Igitur. "Fossilworks: Clidastes". fossilworks.org. Kaddumi, H.F. (2006). "A new genus and species of gigantic marine turtles (Chelonioidea: Cheloniidae) from the Maastrichtian of the Harrana Fauna-Jordan" (PDF). Vertebrate Paleontology. 3 (1). Aaron R. H. Leblanc, Michael W. Caldwell and Nathalie Bardet (2012). "A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics". Journal of Vertebrate Paleontology. 32 (1): 82-104. Kiernan, C.R. 1992. Clidastes Cope, 1868 (Reptilia, Sauria):proposed designation of Clidastes propython Cope, 1869 as the type species. Bulletin of Zoological Nomenclature 49:137-139. ICZN Opinion 1750. 1993. Clidastes Cope, 1868 (Reptilia, Sauria):C. propython Cope, 1869 designated as the type species. Bulletin of Zoological Nomenclature 50: 297. Joshua R. Lively (2019). "Taxonomy and historical inertia: Clidastes (Squamata: Mosasauridae) as a case study of problematic paleobiological taxonomy". Alcheringa: An Australasian Journal of Palaeontology. in press. Callison, G. (1967). Intracranial mobility in Kansas mosasaurs. Lawrence. Charles H. Sternberg Transactions of the Kansas Academy of Science Vol. 30 (Apr. 18, 1919 - Feb. 19, 1921), pp. 119-120. Cope, E.D. 1868. On new species of extinct reptiles. Proceedings of the Academy of Natural Sciences of Philadelphia 20: 181. Dobie, J. L., Daniel, R. W., & Bell, G. L. (June 19, 1986). A Unique Sacroiliac Contact in Mosasaurs (Sauria, Varanoidea, Mosasauridae). Journal of Vertebrate Paleontology, 6, 2, 197-199. Kiernan, C. R. (January 1, 2002). Stratigraphic distribution and habitat segregation of mosasaurs in the Upper Cretaceous of western and central Alabama, with an historical review of Alabama mosasaur discoveries. Journal of Vertebrate Paleontology, 22, 1, 91-103. Lindgren, J & Schulp, A. (September 1, 2010). New material of Prognathodon (Squamata: Mosasauridae), and the mosasaur assemblage of the Maastrichtian of California, U.S.A. Journal of Vertebrate Paleontology, 30, 5.) Wright, K. R. (September 23, 1988). The First Record of Clidastes liodontus (Squamata, Mosasauridae) from the Eastern United States. Journal of Vertebrate Paleontology, 8, 3, 343-345.
Data: 10/02/1997
Emissione: Fauna marina atlantica del Cretaceo
Stato: Tristan da Cunha Nota: Emesso in un foglietto di 4 v. diversi